
Nya insikter om adaptiva reaktioner av det autonoma nervsystemet
STEPHEN W. PORGES, PhD
Den polyvagala teorin beskriver ett autonomt nervsystem som påverkas av det centrala nervsystemet, känsligt för afferenta influenser, kännetecknat av en adaptiv reaktivitet beroende på fylogenin i de neurala kretsarna, och interaktivt med källkärnor i hjärnstammen som reglerar de tvärstrimmiga musklerna i hjärnan. ansikte och huvud. Teorin är beroende av ackumulerad kunskap som beskriver de fylogenetiska övergångarna i det autonoma nervsystemet hos ryggradsdjur. Dess specifika fokus ligger på det fylogenetiska skiftet mellan reptiler och däggdjur som resulterade i specifika förändringar i de vagalbanor som reglerar hjärtat. När källkärnorna för de primära vagala efferenta vägarna som reglerar hjärtat skiftade från den dorsalmotoriska kärnan i vagus hos reptiler till nucleus ambiguus hos däggdjur, utvecklades en ansikte-hjärta-koppling med framväxande egenskaper hos ett socialt engagemangssystem som skulle möjliggöra sociala interaktioner för att reglera det viscerala tillståndet.
Historiska perspektiv på det autonoma nervsystemet
Centrala nervsystemets reglering av viscerala organ är i fokus för flera historiska publikationer som har format strukturen för fysiologisk undersökning. Till exempel, 1872 erkände Darwin det dynamiska neurala förhållandet mellan hjärtat och hjärnan:
”. . .när hjärtat påverkas reagerar det på hjärnan; och hjärnans tillstånd reagerar återigen genom den pneumo-gastriska [vagus] nerven på hjärtat; så att det under varje spänning kommer att finnas mycket ömsesidigt agerande och reaktion mellan dessa, de två viktigaste organen i kroppen.”
Även om Darwin erkände den dubbelriktade kommunikationen mellan inälvorna och hjärnan, minimerade efterföljande formell beskrivning av det autonoma nervsystemet (t.ex. av Langley) betydelsen av centrala regulatoriska strukturer och afferenter. Efter Langley tenderade medicinsk och fysiologisk forskning att fokusera på de perifera motoriska nerverna i det autonoma nervsystemet, med en konceptuell betoning på den parade antagonismen mellan sympatiska och parasympatiska efferenta vägar på målorganen. Detta fokus minimerade intresset för både afferenta vägar och hjärnstammen som reglerar specifika efferenta vägar.
Den tidiga konceptualiseringen av vagus fokuserade på en odifferentierad efferent väg som antogs modulera ”tonen” samtidigt till flera målorgan. Hjärnstamområdena som reglerar den supradiafragma (t.ex. myeliniserade vagala banor som har sitt ursprung i nucleus ambiguus och som slutar i första hand ovanför diafragman) skiljdes inte funktionellt från de som reglerar subdiafragma (t.ex. omyeliniserade vagala banor med ursprung i den dorsalmotoriska vaggans kärna och den dorsalmotoriska vaggan) slutar främst under diafragman). Utan denna distinktion fokuserade forskning och teori på den parade antagonismen mellan den parasympatiska och sympatiska innerveringen till målorgan. Konsekvensen av en betoning på parad antagonism var en acceptans inom fysiologi och medicin av globala konstruktioner som autonom balans, sympatisk tonus och vagal ton.
För mer än 50 år sedan föreslog Hess att det autonoma nervsystemet inte enbart var vegetativt och automatiskt utan istället var ett integrerat system med både perifera och centrala neuroner.3 Genom att betona de centrala mekanismer som förmedlar den dynamiska regleringen av perifera organ, förutsåg Hess att behov av teknik för att kontinuerligt övervaka perifera och centrala neurala kretsar som är involverade i regleringen av visceral funktion.
Den Vagala paradoxen
1992 föreslog jag att en uppskattning av vagal tonus, härledd från mätning av respiratorisk sinusarytmi, skulle kunna användas i klinisk medicin som ett index för stresssårbarhet.4 Istället för att använda de beskrivande måtten på hjärtfrekvensvariabilitet (dvs. slag-till- slagvariabilitet) som ofta används inom obstetrik och pediatrik, betonade tidningen att respiratorisk sinusarytmi har ett neuralt ursprung och representerar det toniska funktionella utflödet från vagus till hjärtat (dvs. hjärtvagal tonus). Således föreslogs det att andningssinusarytmi skulle ge ett mer känsligt index för hälsotillstånd än ett mer globalt mått på hjärtfrekvensvariabilitet från slag till slag som återspeglar obestämda neurala och icke-neurala mekanismer. Uppsatsen presenterade ett kvantitativt tillvägagångssätt som tillämpade tidsserieanalyser för att extrahera amplituden av respiratorisk sinusarytmi som ett mer exakt index för vagal aktivitet. Artikeln tillhandahöll data som visar att friska fullgångna spädbarn hade respiratorisk sinusarytmi med betydligt större amplitud än för tidigt födda barn. Denna idé att använda hjärtfrekvensmönster för att indexera vagal aktivitet var inte ny, efter att ha rapporterats så tidigt som 1910 av Hering.5 Dessutom har samtida studier på ett tillförlitligt sätt rapporterat att vagal blockad via atropin dämpar respiratorisk sinusarytmi hos däggdjur.
Som svar på den här artikeln4 fick jag ett brev från en neonatolog som skrev att han som läkarstudent lärde sig att vagal ton kunde vara dödlig. Han hävdade att kanske för mycket av det goda (dvs vagal ton) kunde vara dåligt. Han syftade naturligtvis på den kliniska risken för neurogen bradykardi. Bradykardi, när det observeras under förlossningen, kan vara en indikator på fosterbesvär. På samma sätt är bradykardi och apné viktiga riskindikatorer för det nyfödda barnet.
Mina kollegor och jag undersökte ytterligare denna förbryllande observation genom att studera det mänskliga fostret under förlossningen. Vi observerade att fetal bradykardi endast inträffade när respiratorisk sinusarytmi var deprimerad (dvs. en andningsrytm i fostrets hjärtfrekvens kan observeras även i frånvaro av de stora bröstväggrörelserna i samband med andning som inträffar efter förlossningen).8 Detta väckte frågan om hur vagala mekanismer kan förmedla både respiratorisk sinusarytmi och bradykardi, eftersom den ena är skyddande och den andra är potentiellt dödlig. Denna inkonsekvens blev den ”vagala paradoxen” och fungerade som motivet bakom den polyvagala teorin.
När det gäller de mekanismer som förmedlar bradykardi och hjärtfrekvensvariationer finns det en uppenbar inkonsekvens mellan data och fysiologiska antaganden. Fysiologiska modeller förutsätter vagal reglering av både kronotropisk kontroll av hjärtat (dvs hjärtfrekvens) och amplituden av respiratorisk sinusarytmi.9,10 Till exempel har det tillförlitligt rapporterats att vagala kardio-hämmande fibrer till hjärtat har konsekventa funktionella egenskaper kännetecknas av bradykardi till neural stimulering och en andningsrytm.9 Men även om det finns situationer där båda åtgärderna samvarierar (t.ex. under träning och kolinerg blockad), finns det andra situationer där åtgärderna tycks återspegla oberoende källor till neural kontroll ( t.ex. bradykardiska episoder associerade med hypoxi, vasovagal synkope och fosterbesvär). I motsats till dessa observerbara fenomen fortsätter forskare att argumentera för en samvariation mellan dessa två parametrar. Denna inkonsekvens, baserad på ett antagande om en enda central vagal källa, är vad jag har kallat vagala paradoxen.
Den polyvagala teorin: tre folygenetiska svarsystem
Undersökning av fylogenin i det autonoma nervsystemet hos ryggradsdjur ger ett svar på den vagala paradoxen. Forskning inom jämförande neuroanatomi och neurofysiologi har identifierat två grenar av vagus, där varje gren stödjer olika adaptiva funktioner och beteendestrategier. Vagalutflödet till hjärtat från en gren manifesteras i respiratorisk sinusarytmi, och utsignalen från den andra grenen manifesteras i bradykardi och möjligen de långsammare rytmerna i hjärtfrekvensvariabilitet. Även om de långsammare rytmerna har antagits ha ett sympatiskt inflytande, blockeras de av atropin.
Den polyvagala teorin7,11–15 artikulerar hur vart och ett av tre fylogenetiska stadier i utvecklingen av det autonoma nervsystemet hos ryggradsdjur är associerat med ett distinkt autonomt subsystem som bibehålls och uttrycks i däggdjur. Dessa autonoma delsystem är fylogenetiskt ordnade och beteendemässigt kopplade till social kommunikation (t.ex. ansiktsuttryck, vokalisering, lyssnande), mobilisering (t.ex. kamp-flykt-beteenden) och immobilisering (t.ex. låtsasdöd, vasovagal synkope och beteendeavstängning).
Det sociala kommunikationssystemet (dvs. systemet för socialt engagemang; se nedan) involverar den myeliniserade vagus, som tjänar till att främja lugna beteendetillstånd genom att hämma sympatiska influenser till hjärtat och dämpa hypotalamus-hypofys-binjureaxeln (HPA).16 Mobiliseringssystemet är beroende av det sympatiska nervsystemets funktion. Den mest fylogenetiskt primitiva komponenten, immobiliseringssystemet, är beroende av den omyeliniserade vagus, som delas med de flesta ryggradsdjur. Med ökad neural komplexitet till följd av fylogenetisk utveckling, berikas organismens beteendemässiga och affektiva repertoar. De tre kretsarna kan konceptualiseras som dynamiska, vilket ger adaptiva svar på säkra, farliga och livshotande händelser och sammanhang.
Endast däggdjur har en myeliniserad vagus. Till skillnad från den omyeliniserade vagus, som har sitt ursprung i den dorsala motoriska kärnan i vagus med pre- och postganglioniska muskarina receptorer, har den myeliniserade däggdjursvagus sitt ursprung i nucleus ambiguus och har preganglioniska nikotinreceptorer och postganglioniska muskarina receptorer. Den omyeliniserade vagus delas med andra ryggradsdjur, inklusive reptiler, amfibier, teleostar och elasmobranchs.
Vi undersöker nu möjligheten att extrahera olika funktioner i hjärtfrekvensmönstret för att dynamiskt övervaka de två vagala systemen. Preliminära studier i vårt laboratorium stödjer denna möjlighet. I dessa studier har vi blockerat de nikotiniska preganglionreceptorerna med hexametonium och de muskarina receptorerna med atropin. Uppgifterna samlades in från präriesorken17 som har en mycket hög omgivande vagal ton. Dessa preliminära data visade att nikotinblockad hos flera djur selektivt tar bort respiratorisk sinusarytmi utan att dämpa amplituden av de lägre frekvenserna i hjärtfrekvensvariabilitet. Däremot tar blockering av de muskarina receptorerna med atropin bort både de låga och andningsfrekvenserna.
Konsistent med Jacksonisk upplösning?
De tre kretsarna är organiserade och svarar på utmaningar i en fylogenetiskt bestämd hierarki som överensstämmer med Jacksons princip om upplösning. Jackson föreslog att i hjärnan hämmar högre (dvs. fylogenetiskt nyare) neurala kretsar lägre (dvs. fylogenetiskt äldre) neurala kretsar och ”när de högre plötsligt görs funktionslösa, desto lägre ökning i aktivitet.”18 Även om Jackson föreslog upplösning för att förklara. förändringar i hjärnans funktion på grund av skador och sjukdomar, den polyvagala teorin föreslår en liknande fylogenetiskt ordnad hierarkisk modell för att beskriva sekvensen av autonoma svarsstrategier på utmaningar.
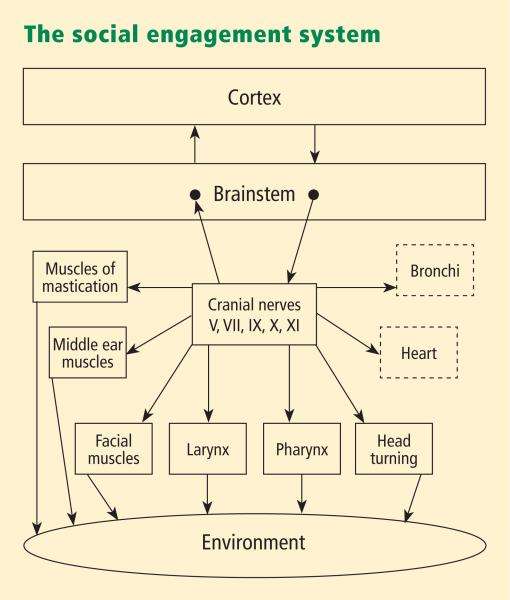
Funktionellt, när miljön upplevs som säker, uttrycks två viktiga egenskaper. För det första regleras det kroppsliga tillståndet på ett effektivt sätt för att främja tillväxt och restaurering (t.ex. visceral homeostas). Detta görs genom ett ökat inflytande från däggdjurs myeliniserade vagalmotoriska vägar på pacemakern som saktar ner hjärtat, hämmar kamp-flyg-mekanismerna i det sympatiska nervsystemet, dämpar stressresponssystemet i HPA-axeln (t.ex. kortisol) och minskar inflammation genom att modulera immunreaktioner (t.ex. cytokiner). För det andra, genom evolutionsprocessen, blev hjärnstammens kärnor som reglerar den myeliniserade vagus integrerade med kärnorna som reglerar musklerna i ansiktet och huvudet. Denna länk resulterar i den dubbelriktade kopplingen mellan spontana beteenden för socialt engagemang och kroppsliga tillstånd. Specifikt uppstod ett integrerat socialt engagemangssystem hos däggdjur när den neurala regleringen av viscerala tillstånd som främjar tillväxt och restaurering (via den myeliniserade vagus) kopplades neuroanatomiskt och neurofysiologiskt till den neurala regleringen av musklerna som kontrollerar ögonblick, ansiktsuttryck, lyssnande och prosodi.
Det mänskliga nervsystemet, liknande det hos andra däggdjur, utvecklades inte bara för att överleva i säkra miljöer utan också för att främja överlevnad i farliga och livshotande sammanhang. För att uppnå denna adaptiva flexibilitet behöll det mänskliga nervsystemet ytterligare två primitiva neurala kretsar för att reglera defensiva strategier (dvs. kamp-flykt och dödslåtsas beteenden). Det är viktigt att notera att socialt beteende, social kommunikation och visceral homeostas är oförenliga med de neurofysiologiska tillstånd och beteenden som främjas av de två neurala kretsar som stöder försvarsstrategier. Således, via evolution, behåller det mänskliga nervsystemet tre neurala kretsar, som är i en fylogenetiskt organiserad hierarki. I denna hierarki av adaptiva svar används den senaste kretsen först; om den kretsen inte ger säkerhet, rekryteras de äldre kretsarna sekventiellt.
Undersökning av fylogenin för reglering av ryggradsdjurshjärtat har lett till utvinning av fyra principer som ger en grund för testning av hypoteser som relaterar specifika neurala mekanismer till socialt engagemang, kamp-flykt och dödslåtsasbeteenden:
Det sker ett fylogenetiskt skifte i regleringen av hjärtat från endokrin kommunikation till omyeliniserade nerver och slutligen till myeliniserade nerver.
Det finns en utveckling av motsatta neurala mekanismer för excitation och hämning för att ge snabb reglering av graderad metabolisk produktion.
En ansikte-hjärta-förbindelse utvecklades när källkärnor för vagala banor skiftade ventralt från den äldre dorsalmotoriska kärnan till nucleus ambiguus. Detta resulterade i en anatomisk och neurofysiologisk koppling mellan neural reglering av hjärtat via den myeliniserade vagus och de speciella viscerala efferenta banorna som reglerar de tvärstrimmiga musklerna i ansiktet och huvudet, vilket bildar ett integrerat socialt engagemangssystem.
Med ökad kortikal utveckling uppvisar cortex större kontroll över hjärnstammen via direkta (t.ex. corticobulbar) och indirekta (t.ex. korticoretikulära) neurala banor som har sitt ursprung i motorisk cortex och som slutar i källkärnorna för de myeliniserade motoriska nerverna som kommer ut från hjärnstammen (t.ex. , specifika neurala banor inbäddade i kranialnerverna V, VII, IX, X och XI), som kontrollerar visceromotoriska strukturer (dvs hjärta, bronkier) såväl som somato-motoriska strukturer (muskler i ansikte och huvud).
NEUROCEPTION: Kontextuell cueing av adaptiva, maladaptiva fysiologiska tillstånd
För att effektivt byta från defensiva till sociala engagemangsstrategier måste däggdjursnervsystemet utföra två viktiga adaptiva uppgifter: (1) bedöma risker, och (2) om miljön upplevs som säker, hämma de mer primitiva limbiska strukturerna som styr kampen, flykt eller frysningsbeteenden.
Varje stimulans som har potential att öka en organisms upplevelse av säkerhet har potentialen att rekrytera de evolutionärt mer avancerade neurala kretsar som stödjer det sociala engagemangssystemets prosociala beteenden.
Nervsystemet, genom bearbetning av sensorisk information från omgivningen och från inälvorna, utvärderar kontinuerligt risker. Eftersom den neurala utvärderingen av risk inte kräver medveten medvetenhet och kan involvera subkortikala limbiska strukturer21 introducerades termen neuroception22 för att betona en neural process, skild från perception, som kan särskilja miljömässiga (och viscerala) egenskaper som är säkra, farliga , eller livshotande. I säkra miljöer regleras det autonoma tillståndet adaptivt för att dämpa sympatisk aktivering och för att skydda det syreberoende centrala nervsystemet, särskilt cortex, från de metaboliskt konservativa reaktionerna från det dorsala vagala komplexet. Men hur vet nervsystemet när miljön är säker, farlig eller livshotande, och vilka neurala mekanismer utvärderar denna risk?
Miljökomponenter av neuroception
Neuroception representerar en neural process som gör det möjligt för människor och andra däggdjur att engagera sig i sociala beteenden genom att skilja säkert från farliga sammanhang. Neuroception föreslås som en rimlig mekanism som förmedlar både uttrycket och störningen av positivt socialt beteende, känsloreglering och visceral homeostas.7,22 Neuroception kan triggas av funktionsdetektorer som involverar områden av temporal cortex som kommunicerar med amygdalas centrala kärna och den periakveduktala gråa, eftersom limbisk reaktivitet moduleras av temporala cortex-svar på avsikten med röster, ansikten och handrörelser. Således översätts neuroceptionen hos bekanta individer och individer med lämpligt prosodiska röster och varma, uttrycksfulla ansikten till en social interaktion som främjar en känsla av säkerhet.
Hos de flesta individer (dvs de utan en psykiatrisk störning eller neuropatologi) utvärderar nervsystemet risk och matchar neurofysiologiskt tillstånd med den faktiska risken i miljön. När miljön bedöms som säker, hämmas de defensiva limbiska strukturerna, vilket möjliggör socialt engagemang och lugna viscerala tillstånd att uppstå. Däremot upplever vissa individer en obalans och nervsystemet bedömer miljön som farlig även när den är säker. Denna oöverensstämmelse resulterar i fysiologiska tillstånd som stödjer strids-, flykt- eller frysbeteenden, men inte beteenden för socialt engagemang. Enligt teorin kan social kommunikation uttryckas effektivt genom det sociala engagemangssystemet endast när dessa defensiva kretsar hämmas.
Andra bidragsgivare till neuroception
Riskdragen i miljön driver inte bara neuroceptionen. Afferent feedback från inälvorna ger en viktig förmedlare av tillgängligheten för prosociala kretsar som är förknippade med beteenden för socialt engagemang. Till exempel förutspår den polyvagala teorin att tillstånd av mobilisering skulle äventyra vår förmåga att upptäcka positiva sociala signaler. Funktionellt sett färgar viscerala tillstånd vår uppfattning om föremål och andra. Således kan samma egenskaper hos en person som engagerar en annan resultera i en rad olika resultat, beroende på målindividens fysiologiska tillstånd. Om personen som är engagerad befinner sig i ett tillstånd där systemet för socialt engagemang är lättillgängligt, kommer sannolikt de ömsesidiga prosociala interaktionerna att inträffa. Men om individen befinner sig i ett tillstånd av mobilisering, kan samma engagerande svar besvaras med de asociala dragen som tillbakadragande eller aggression. I ett sådant tillstånd kan det vara mycket svårt att dämpa mobiliseringskretsen och göra det möjligt för systemet för socialt engagemang att komma tillbaka på nätet.
Insulan kan vara involverad i förmedlingen av neuroception, eftersom den har föreslagits som en hjärnstruktur involverad i att förmedla den diffusa återkopplingen från inälvorna till kognitiv medvetenhet. Funktionella avbildningsexperiment har visat att insula spelar en viktig roll i upplevelsen av smärta och upplevelsen av flera känslor, inklusive ilska, rädsla, avsky, lycka och sorg. Critchley föreslår att interna kroppstillstånd är representerade i insula och bidrar till tillstånd av subjektiv känsla, och han har visat att aktivitet i insula korrelerar med interoceptiv noggrannhet.
Summarum
Den polyvagala teorin föreslår att utvecklingen av det autonoma nervsystemet hos däggdjur tillhandahåller de neurofysiologiska substraten för adaptiva beteendestrategier. Den föreslår vidare att fysiologiskt tillstånd begränsar utbudet av beteende och psykologisk erfarenhet. Teorin kopplar utvecklingen av det autonoma nervsystemet till affektiv upplevelse, känslomässiga uttryck, ansiktsgester, röstkommunikation och tillfälligt socialt beteende. På detta sätt ger teorin en rimlig förklaring till den rapporterade samvariationen mellan atypisk autonom reglering (t.ex. minskad vagal och ökad sympatisk påverkan på hjärtat) och psykiatriska och beteendemässiga störningar som innebär svårigheter att reglera lämpliga sociala, emotionella och kommunikationsbeteenden.
Den polyvagala teorin ger flera insikter om det fysiologiska tillståndets adaptiva natur. För det första betonar teorin att fysiologiska tillstånd stödjer olika klasser av beteende. Till exempel skulle ett fysiologiskt tillstånd som kännetecknas av ett vagalt tillbakadragande stödja mobiliseringsbeteendet av kamp och flykt. Däremot skulle ett fysiologiskt tillstånd som kännetecknas av ökat vagalt inflytande på hjärtat (via myeliniserade vagala vägar med ursprung i nucleus ambiguus) stödja spontana beteenden för socialt engagemang. För det andra betonar teorin bildandet av ett integrerat socialt engagemangssystem genom funktionella och strukturella länkar mellan neural kontroll av de tvärstrimmiga musklerna i ansiktet och de glatta musklerna i inälvorna. För det tredje föreslår den polyvagala teorin en mekanism – neuroception – för att utlösa eller hämma försvarsstrategier.
Avslutande ord
Dr. Porges rapporterade att han inte har några ekonomiska intressen eller relationer som utgör en potentiell intressekonflikt med denna artikel. Förberedelsen av detta manuskript stöddes delvis av ett anslag från National Institutes of Health.
Artikeln har hämtats från National Library of Medicine och översatts för publicering här på kroppogestalt.se 2024-01-05 av Thomas Sonefors.
Hela artikeln på engelska med tillhörande referenser finns här.


